Research projects
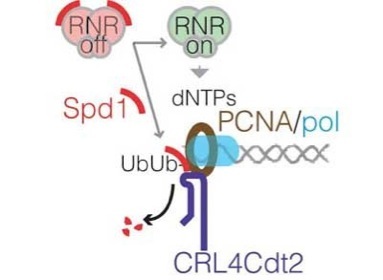
Eukaryotic chromosomes contain many origins at which replication initiates, but DNA replication is tightly regulated to ensure that this does not occur in inappropriate circumstances. For instance, DNA replication must be prevented in non-proliferating cells. When DNA replication is switched on, initiation must occur from origins only once per cell cycle, otherwise amplification of chromosomal regions would occur. This regulation involves proteolysis of factors needed for initiation during S phase, thus preventing re-initiation. Our recent work has shown that a specific ubiquitin ligase, CRL4Cdt2, is involved in this regulation via ubiquitylation and proteolysis of the helicase-loading factor Cdt1, during S phase. Recent work has shown that proteolysis of Cdt1 uses a novel mechanism where a protein at the replication fork itself is necessary for this process. PCNA, a ring shaped molecule involved in holding polymerases on the template, is loaded onto DNA in S phase. This process involves transient opening of the PCNA ring so that it encompasses the DNA double helix. PCNA also interacts with Cdt1, and acts as a platform when DNA associated to allow ubiquitylation of Cdt1 by CRL4Cdt2 , thus synchronising proteolysis with S phase onset. Only DNA-associated PCNA can promote this reaction and we are trying to determine why this is the case.
CRL4Cdt2 also targets Spd1 via the same mechanism in S phase. Spd1 is an inhibitor of ribonucleotide reductase (RNR), which catalyses the rate limiting step in dNTP production. Thus proteolysis of Spd1 in S phase helps to ensure that the elongation step of DNA replication is efficient.
Cells where CRL4Cdt2 is defective are deficient in DNA repair and this is only partially due to defects in Spd1 proteolysis. We are determining if this is due to stabilisation of Cdt1 or other substrates of CRL4Cdt2.
See Guarino et al. (2011) and Salguero et al. (2012) for more details.
Proteolytic regulation of DNA replication regulators
Regulation of DNA replication in meiosis
DNA replication usually alternates with mitosis and cell division. However, in meiosis, an exception to this occurs, with pre-meiotic S phase being followed by two rounds of nuclear division (meiosis I and meiosis II) without intervening chromosome replication. This allows haploid cells to generated from a diploid progenitor and is an essential feature of sexual reproduction. We are investigating how DNA replication is repressed between the meiotic nuclear divisions in fission yeast. Work from using a number of organisms has shown that the loading the Mcm2-7 helicase onto DNA is repressed in the meiosis I-II interval. In recent work we have shown that when we force Mcm2-7 loading onto DNA during this interval it only results in a partial round of DNA replication, and inefficient elongation appears to contribute to the block to DNA replication during this interval. We are currently investigating why the elongation step of DNA replication is inefficient and whether this represents an important ‘fail-safe’ mechanism to ensure that meiotic progeny are haploid with no re-replicated chromosomal regions.
See Hua et al. (2013) for more details.
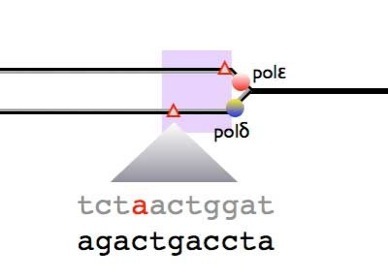
Analysis of ‘cancer’ mutations in DNA replication enzymes
Defects in DNA repair mechanism are well known to be associated with cancer predisposition syndromes. Evidence from mice also suggests that DNA replication defects cause cancer but it has been until recently less clear whether this is important in human cancer. Recently, the Tomlinson group (Wellcome Institute for Human Genetics, Oxford) has shown that mutations in the exonuclease domain of replicative polymerases are associated with familial predisposition to colorectal cancer (Palles et al. 2012). In collaboration with this group, we have modelled these mutations in fission yeast, and shown them to have a mutator phenotype. This exciting finding suggests that subtle defects in replicative enzymes can be one source of the genetic instability that leads to cancer development.
See Palles et al. (2012) for more details.