Mark Howarth lab
\
Our website has moved to http://www.bioch.ox.ac.uk/howarth/index.htm
Please update your bookmarks.
You will be automatically redirected.
Copyright © 2014 Mark Howarth. All rights reserved.


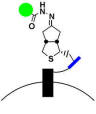 Mark Howarth lab
Mark Howarth lab