Figure 8
|
|
Jarvis Lab |
Figure 8 |
|
|
|
|
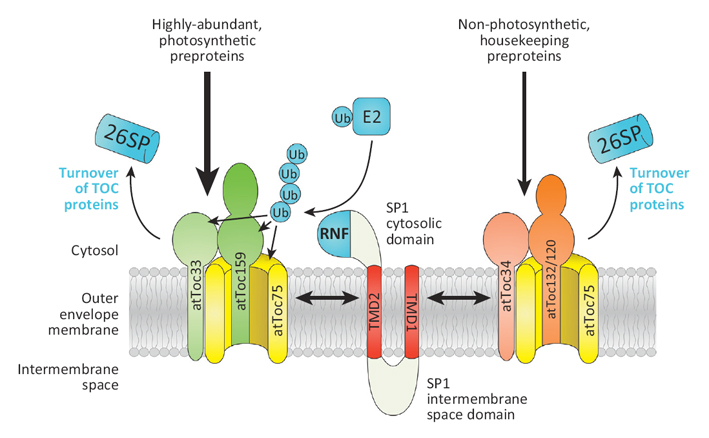
Figure 8: The Role of SP1 in the Regulation of Plastid Protein Import |
|
The SP1 protein is held in the plastid outer envelope membrane by two transmembrane domains (TMD1, TMD2). Its RING finger (RNF) domain is oriented towards the cytosol where it recruits E2 ubiquitin (Ub) conjugating enzyme to mediate the ubiquitination of TOC (translocase of the outer chloroplast membrane) proteins, which are recognized by the SP1 intermembrane-space domain. Once ubiquitinated, TOC components are degraded by the cytosolic 26S proteasome (26SP). In Arabidopsis (and other plants), components of the TOC machinery exist in different isoforms (e.g., atToc33 and atToc34), and this enables the formation of different TOC complexes with different client specificities: the main isoforms (those that predominate in chloroplasts) form TOC complexes with specificity for precursors of the photosynthetic apparatus, whereas other isoforms form complexes with specificity for ‘housekeeping’ precursor proteins. The balance of the import pathways mediated by these different TOC complexes controls the composition of the organellar proteome, and this in turn influences the developmental fate and functions of the organelle. The SP1 protein plays a critical role in controlling this balance by facilitating the turnover of TOC components and enabling reorganization of the import machinery. Taken from Trends Cell Biol., 2013, 23: 399-408. |
|
|
Last updated: Dec 2023
Paul Jarvis